

Molecular Endocrinology and Oncology Research Centre, Department of Nutrition, Université Laval Medical Research Centre and Université Laval
andre.tchernof@crchul.ulaval.ca

Numerous studies have shown the strong and independent association between fat accumulation on anatomical structures such as the mesentery and greater omentum (i.e., visceral fat accumulation) and risk factors for type 2 diabetes and cardiovascular disease [1]. This strong association suggests a close physiological link between fat cells located within the visceral fat compartments, the visceral adipocytes, and metabolic abnormalities. This brief review will discuss potential mechanisms relating the visceral adipocyte to cardiometabolic risk factors.
 [Click to enlarge]
[Click to enlarge]
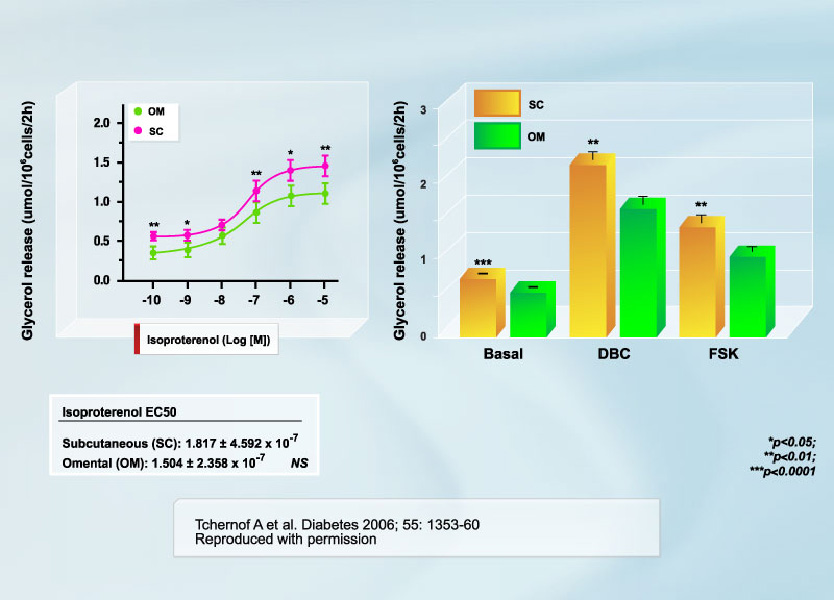
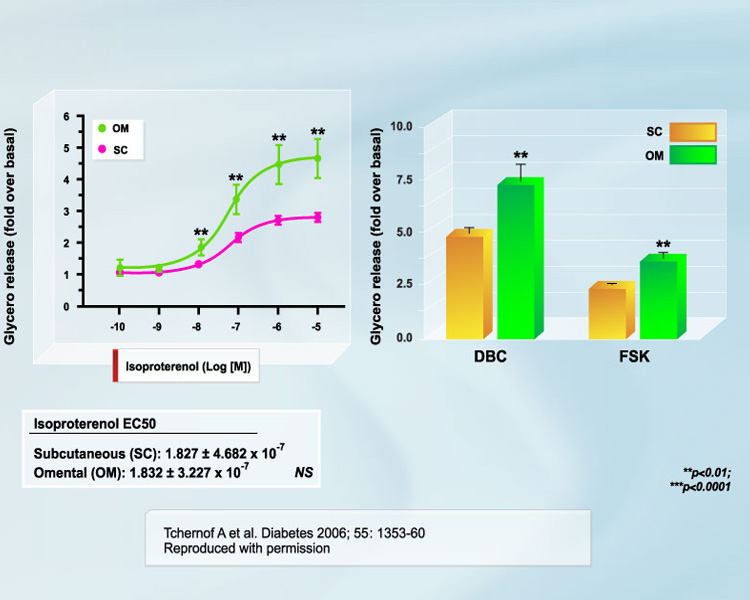
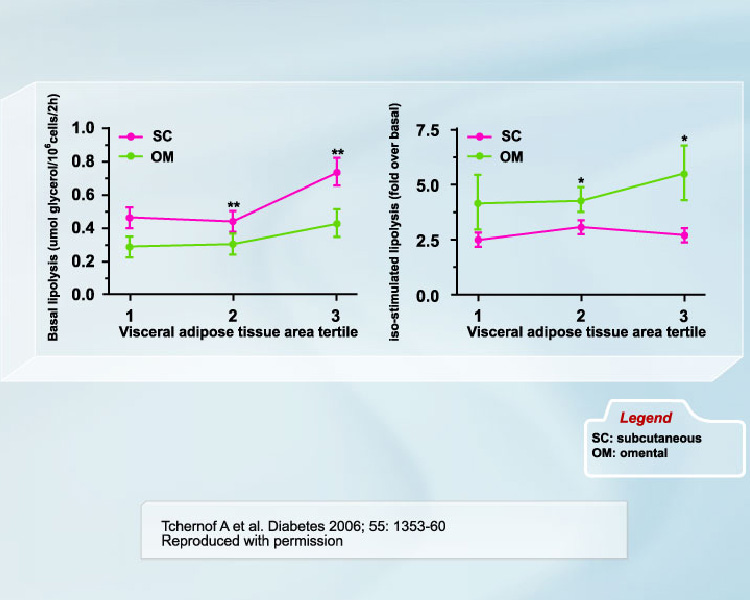
The hypertriglyceridemic state of visceral obesity is primarily due to VLDL overproduction [2, 3]. Availability of fatty acids in the liver is recognized as the primary determinant of this overproduction [3], which has led to the hypothesis that an increased fatty acid flux from adipose tissue located within the abdominal cavity through the portal vein to the liver could potentially explain visceral obesity-related hypertriglyceridemia [4]. Visceral adipose cells are believed to be hyperlipolytic and poorly responsive to insulin inhibition [4-6]. A recent study by our group is the largest to date to be performed in women on this issue [7]. We found that although lipolytic rates were higher in subcutaneous adipocytes when values were expressed on an absolute basis (per cell) (Figure 1), the responsiveness of omental adipocytes to positive lipolytic stimuli was much higher than that of subcutaneous adipocytes (Figure 2). We also reported in the study that omental vs. subcutaneous differences in lipolysis were relatively constant throughout the spectrum of adiposity values (Figure 3). Conversely, in men, no difference was observed in the size of omental vs. subcutaneous fat cells. Accordingly, basal lipolysis and lipolytic responses to positive stimuli expressed either as a function of cell number or as fold response over basal levels were not significantly different in omental vs. subcutaneous fat cells [8]. We suggest that regional differences in adipocyte size (or lack thereof) are important determinants of regional differences in adipose tissue metabolism.
 [Click to enlarge]
[Click to enlarge]
Other mechanisms could also explain the close association between excess visceral fat accumulation and cardiometabolic risk factors. Results on ectopic fat accumulation have led to the suggestion that insulin resistance may be due to increased lipid burden on skeletal muscle and liver from a reduced capacity for excess fat storage in the presence of excess energy intake [9-11]. According to this hypothesis, insulin resistance could be due not only to lipids released from fat, but also to a reduced capacity for excess lipid handling and storage in peripheral fat depots [11, 12].
 [Click to enlarge]
[Click to enlarge]
More recently, the endocrine and paracrine nature of the adipose organ (Figure 4) has emerged as a new line of investigation, and adipose tissue-secreted cytokines (adipokines) are believed to mediate part of the link between visceral fat accumulation and cardiometabolic risk. Studies in humans and rodents have shown that the various cell types found in visceral adipose tissue (adipocytes, stromal or vascular cells, macrophages, etc.) release cytokines such as resistin, adiponectin, TNF-a, and IL-6, which may contribute to obesity-related insulin resistance [13-15].
 [Click to enlarge]
[Click to enlarge]
Adiponectin, a cytokine produced in adipose tissue, has been shown to be decreased in obesity, type 2 diabetes, and atherosclerosis [16, 17]. Recent data indirectly suggest that adiponectin is reduced primarily in omental fat of obese subjects, which would indicate yet another possible preferential link between visceral fat accumulation and cardiometabolic risk factors. Taken together, visceral adipose tissue fatty acid release, reduced peripheral lipid storage, and secreted cytokines may all be involved in the etiology of insulin resistance, the metabolic syndrome, and related global cardiometabolic risk.
References
- Després JP and Lemieux I. Abdominal obesity and metabolic syndrome. Nature 2006; 444: 881-7
- Lewis GF, Carpentier A, Adeli K, et al. Disordered fat storage and mobilization in the pathogenesis of insulin resistance and type 2 diabetes. Endocr Rev 2002; 23: 201-29
- Lewis GF. Fatty acid regulation of very low density lipoprotein production. Curr Opin Lipidol 1997; 8: 146-53.
- Bjorntorp P. "Portal" adipose tissue as a generator of risk factors for cardiovascular disease and diabetes. Arteriosclerosis 1990; 10: 493-6
- Kissebah AH, Freedman DS and Peiris AN. Health risks of obesity. Med Clin North Am 1989; 73: 111-38.
- Kissebah AH and Peiris AN. Biology of regional body fat distribution: relationship to non-insulin-dependent diabetes mellitus. Diabetes/Metabolism Reviews 1989; 5: 83-109.
- Tchernof A, Belanger C, Morisset AS, et al. Regional differences in adipose tissue metabolism in women: minor effect of obesity and body fat distribution. Diabetes 2006; 55: 1353-60.
- Boivin A, Brochu G, Marceau S, et al. Regional differences in adipose tissue metabolism in obese men. Metabolism 2007; 56: 533-40
- Nadler ST and Attie AD. Please pass the chips: genomic insights into obesity and diabetes. J Nutr 2001; 131: 2078-81.
- Reitman ML, Mason MM, Moitra J, et al. Transgenic mice lacking white fat: models for understanding human lipoatrophic diabetes. Ann N Y Acad Sci 1999; 892: 289-96.
- Kim JK, Fillmore JJ, Chen Y, et al. Tissue-specific overexpression of lipoprotein lipase causes tissue-specific insulin resistance. Proc Natl Acad Sci U S A 2001; 98: 7522-7.
- Nadler ST, Stoehr JP, Schueler KL, et al. The expression of adipogenic genes is decreased in obesity and diabetes mellitus. Proc Natl Acad Sci U S A 2000; 97: 11371-6.
- Steppan CM, Bailey ST, Bhat S, et al. The hormone resistin links obesity to diabetes. Nature 2001; 409: 307-12.
- Spranger J, Kroke A, Mohlig M, et al. Adiponectin and protection against type 2 diabetes mellitus. Lancet 2003; 361: 226-8.
- Fried SK, Bunkin DA and Greenberg AS. Omental and subcutaneous adipose tissues of obese subjects release interleukin-6: depot difference and regulation by glucocorticoid. J Clin Endocrinol Metab 1998; 83: 847-50.
- Hu E, Liang P and Spiegelman BM. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J Biol Chem 1996; 271: 10697-703.
- Hotta K, Funahashi T, Arita Y, et al. Plasma concentrations of a novel, adipose-specific protein, adiponectin, in type 2 diabetic patients. Arterioscler Thromb Vasc Biol 2000; 20: 1595-9.
